
نحو منصة مؤتمتة بالكامل خوارزمية لتصميم النظم الحيوية
نحو منصة مؤتمتة بالكامل خوارزمية لتصميم النظم الحيوية
لتحميل الملف الأصلي: Towards a fully automated algorithm driven platform for biosystems design
محمد حميض رعد ، ران تشاو ، Scott ، Jiazhang Lian ، سوراب سينها، هويمين تشاو
نبذة مختصرة
غالبًا ما يتطلب الحصول على البيانات على نطاق واسع وتحليلها في التنفيذ الناجح لدورة التصميم والبناء والاختبار والتعلم (DBTL) في تصميم النظم الحيوية. ومع ذلك ، فقد تم إعاقة ذلك منذ وقت طويل بسبب التكلفة التجريبية ، والتغيرات ، والتحيزات ، والرؤى الضائعة من طرق التحليل التقليدية. هنا ، نقوم بالإبلاغ عن تطبيق نظام روبوتي متكامل مقترن بخوارزميات التعلم الآلي من أجل أتمتة عملية DBTL لتصميم النظم الحيوية. كدليل على المفهوم ، لقد أثبتنا قدرتها من خلال تحسين مسار الليكوبين الحيوي. يقوم هذا النظام الآلي الآلي بالكامل ، BioAutomata ، بتقييم أقل من 1٪ من المتغيرات المحتملة بينما يتفوق على الفحص العشوائي بنسبة 77٪. نموذج تنبؤي مقترن وخوارزمية بايزية ، حدد التجارب التي يتم إجراؤها بواسطة مسبك إلينوي البيولوجي للصناعات الحيوية المتقدمة (iBioFAB). تتفوق BioAutomata مع مشاكل تحسين الصندوق الأسود ، حيث تكون التجارب باهظة الثمن وصاخبة ولا يعتمد نجاح التجربة على المعرفة المسبقة الشاملة بالآليات البيولوجية.
المقدمة
تم استكشاف النظم البيولوجية مثل البروتينات والمسارات والخلايا بأكملها بشكل متزايد لمجموعة واسعة من تطبيقات التكنولوجيا الحيوية . ومع ذلك ، نظرًا لتعقيد الأنظمة البيولوجية ومكوناتها الكثيرة والعديد من التفاعلات غير المعروفة بينها ، يجب إجراء العديد من جولات التصميم والبناء والاختبار والتعلم (DBTL) . كانت هناك جهود كثيرة للإسراع في دورة DBTL ، والأتمتة الحيوية مثل إلينوي Biological Foundry للتطوير الحيوي المتقدم (iBioFAB) و Edinburgh Genome Foundry كانت قفزة مهمة لا يمكن إنكارها نحو أتمتة مكونات التصميم والبناء والاختبار للدورة . ومع ذلك ، بخلاف بعض التطبيقات المحددة والضيقة، لا يوجد مثال على الأتمتة والتكامل لمكون التعلم لإغلاق دورة DBTL وتمكين تكرار هذه الدورة بأقل تدخل بشري.
علاوة على ذلك ، لا يقتصر التشغيل الآلي على بناء واختبار عناصر الدورة وبالنظر إلى الكم الهائل من البيانات التي تم إنشاؤها بواسطة الأساسيات الحيوية الحديثة ، فإن أتمتة مكون التعلم أمر حاسم أيضًا. تعتبر المساعدة من خوارزميات الكمبيوتر واستخدام النماذج الإحصائية والتعلم الآلي ذات أهمية خاصة بالنظر إلى تعقيد معظم النظم البيولوجية ذات الأهمية العملية والبعد الكبير لمهام التحسين المطلوبة لتقدير مثل هذه الأنظمة ومعالجتها. يمكن تصميم النظم البيولوجية التي تتراوح من البروتينات الفردية إلى المسارات بأكملها باستخدام النماذج الإحصائية وخوارزميات التعلم الآلي وتعلم التعزيز ومجموعة كاملة من النماذج الفيزيائية الحيوية . ومع ذلك ، فقد تركز معظم التقدم المحرز في أتمتة دورة DBTL على أحد عناصر هذه الدورة حيث يمكن أن يؤدي دمج كل هذه المكونات إلى تأثير تآكلي لتمكين كمية كبيرة من البيانات عالية الأبعاد يمكن الحصول عليها وتحليلها بواسطة دورة DBTL مؤتمتة بالكامل.
للتغلب على هذه القيود ، ندمج iBioFAB ، منصة روبوتية مؤتمتة بالكامل وتنوعا 7 مع خوارزمية التعلم الآلي. تصمم منصة BioAutomata هذه التجارب وتنفذها وتحلل البيانات لتحسين العملية البيولوجية المحددة من قِبل المستخدم بطريقة تكرارية. تقوم BioAutomata بتدريب نموذج احتمالي على البيانات التي تم إنشاؤها في البداية (أو المتاحة) وتقرر أفضل النقاط لمساحة التحسين لتقييمها ، أي النقاط التي من المرجح أن تؤدي إلى نظام بيولوجي محسن. يؤدي هذا إلى تقليل إجمالي عدد التجارب اللازمة للعثور على الحد الأقصى لمساحة التحسين. يعتبر إطار التحسين هذا مثاليًا للحالات التي يكون فيها الهدف هو العثور على أوبتيما لوظيفة الصندوق الأسود وحيث يكون الحصول على البيانات باهظ الثمن وصاخبه ، وهذا صحيح في جوهره في تصميم النظم الحيوية. وقد ثبت أن تحسين النظرية الافتراضية أداة قوية في مجالات أخرى مثل هندسة البروتين .
كدليل على المفهوم ، نقوم بتحسين مسار إنتاج اللايكوبين ، أي تحسين التعبير عن الجينات المشاركة في التركيب الحيوي (مدخلات الوظيفة) لتحقيق أعلى إنتاج لليكوبين (مخرجات الوظيفة). تم استخدام الليكوبين بشكل تقليدي كإضافة غذائية وتلوين ، لكن في الآونة الأخيرة ، اقترحت العديد من التقارير آثاره كمضاد للأكسدة ومضاد للسرطان وللوقاية من أمراض القلب والأوعية الدموية . نظرًا للقيمة التجارية المرتفعة للليكوبين ، كان مسار التركيب الحيوي لليكوبين هدفًا للعديد من المساعي الهندسية الأيضية. في حين أن هناك استراتيجيات أخرى مثل حذف أو إفراط في التعبير عن الجينات الداخلية في الكائن الحي لدفع التدفق نحو منتج المسار ، أو ببساطة تحسين ظروف التخمير ، فإن تعبير تعبير الجينات التخليقية الحيوية هو الخيار الأول غالبًا. من خلال الجمع بين خوارزمية التحسين Bayesian ونظام أتمتة iBioFAB ، نقوم بتقييم <1٪ من جميع قيم التعبير القابلة للتوليف المحتملة للجينات المكونة مقابل الإنتاج (منظر تعبير للإنتاج) للعثور على سلالة تنتج عيارًا عاليًا من اللايكوبين. تشير كل نقطة في هذا المشهد إلى كمية إنتاج المادة الكيميائية المطلوبة بالنظر إلى مستوى التعبير المعين لكل جين. بعد التصميم والإعداد المبدئي لهذه BioAutomata ، يتغير دور الباحثين من كونهم المحركين للتجارب إلى المشرفين على النظام بينما تصمم منصة التحسين المعتمدة على الخوارزمية وتجري التجارب لتعظيم الوظيفة الموضوعية التي حددها الباحثون.
النتائج
منصة أوتوماتيكية بالكامل يحركها الخوارزمية BioAutomata
في تصميم النظم الحيوية ، عادة ما يكون إجراء تجارب المختبرات الرطبة باهظًا ، ويستغرق وقتًا طويلًا وعرضة للخطأ. لذلك ، يكون تحسين النظام البيولوجي أكثر فعالية عندما يتم تقليل عدد التجارب التي تم إجراؤها. يظهر النهج المقترح لتحقيق ذلك في الشكل . في هذا السياق ، تتمثل الخطوة الأولى في التحسين في تحديد التصميم الأولي للمدخلات والمخرجات للنظام بالإضافة إلى الوظيفة الموضوعية. بعد الإعداد الأولي ، يجب اختيار نموذج تنبؤي وسياسة الاستحواذ لتقدير المشهد في ضوء البيانات المتاحة حاليًا واختيار النقاط التالية المراد تقييمها والتجارب المراد تنفيذها. بعد اختيار جميع عناصر النظام (التصميم الأولي ، وسياسة الاستحواذ ، والإعداد التجريبي ، والحصول على البيانات ، والنموذج التنبئي) ، يمكن لـ BioAutomata بدء التحسين. أولاً ، تختار سياسة الاستحواذ النقاط المراد تقييمها. بعد ذلك ، يقوم iBioFAB بإجراء التجارب التي تقيم النقاط المحددة لياقتهم البدنية وتعيد البيانات إلى النموذج التنبئي. سيقوم النموذج بعد ذلك بتحديث إيمانه بالمشهد الطبيعي اعتمادًا على البيانات المقدمة حديثًا. أخيرًا ، ستختار سياسة الاستحواذ النقاط التي سيتم تقييمها بعد ذلك مع توجيه النموذج التنبؤي المحدث.
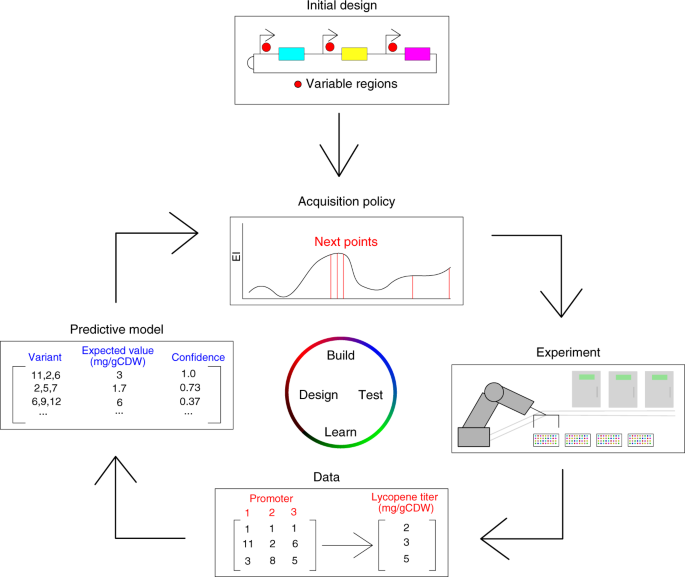
سير العمل العام ل BioAutomata. بعد تعيين المعلمات الأولية ، وتصميم مساحة التسلسل للمناطق المتغيرة (مثل المتغيرات المروجية في مجموعة مسار مسار اندماجي) ، وتحديد الوظيفة الهدف ، تختار BioAutomata التجارب التي من المتوقع أن تؤدي إلى أفضل تحسين للعائد ، وتنفذ تلك التجارب ، يولد البيانات ويتعلم منها ، وتحديث نموذجها التنبئي في ضوء الأدلة المقدمة حديثا. بعد ذلك ستقرر التجارب التالية التي يتعين القيام بها للوصول إلى الهدف الذي حدده المستخدم أثناء محاولة تقليل عدد التجارب وتكلفة المشروع.
تحديد النموذج التنبؤي وسياسة الاستحواذ
نظرًا لأن الهدف هو إيجاد الحد الأقصى لوظيفة الصندوق الأسود حيث يكون الحصول على البيانات باهظ الثمن وصاخبًا ، فقد سعينا إلى استخدام التحسين Bayesian ، والذي يعد مثاليًا لحل هذه المشكلات.تحسين Bayesian هو أسلوب قوي أثبت أنه يتفوق على العديد من الخوارزميات في تحسين هذه الوظائف الصعبة . باختصار ، يقوم بإنشاء نموذج احتمالي ويستخدم هذا النموذج لاتخاذ قرارات بشأن مكان التقييم التالي لتعظيم التقدم المتوقع المحرز مع كل تقييم للوظيفة وبالتالي تقليل عدد التقييمات ، أي التجارب المطلوبة للعثور على الحد الأقصى. تأخذ الخوارزمية النتيجة المتوقعة لكل تقييم وكذلك الثقة في هذه النتيجة المتوقعة في الاعتبار. لاستخدام هذه الخوارزمية ، يجب اختيار وظيفتين رئيسيتين ، نموذج احتمالي لوضع افتراضات حول المشهد في ضوء البيانات المتاحة وسياسة الاستحواذ لاقتراح النقطة التي يجب تقييمها بعد ذلك لتعظيم التقدم المتوقع نحو الحد الأقصى.
استخدمنا عملية Gaussian (GP) كنموذج تنبؤي لتعيين قيمة ومستوى ثقة متوقعين لجميع النقاط التي لم يتم تقييمها. تم اختيار GP بسبب مرونته وتطبيقاته الواسعة . يعين GP متوسطًا وتباينًا لكل نقطة في المنظر الطبيعي ومع تقييم المزيد من النقاط ، يتم تحديث الوسط والتباين وفقًا لذلك (الشكل التكميلي ).
تدفع وظيفة الاستحواذ الاتجاه التجريبي إلى تحقيق التقدم المتوقع نحو المستوى الأمثل. بالنظر إلى القيمة المتوقعة والثقة في هذه القيمة ، نواجه مفاضلة بين الاستكشاف والاستغلال. إذا كانت النقاط التي تم اختبارها هي تلك التي تمتلك أكبر القيم المتوقعة ، فإننا نجازف فقط بإيجاد الحد الأقصى المحلي. وبالتالي ، نريد استكشاف المزيد (التركيز على النقاط التي يكون النموذج غير مؤكد فيها). ومع ذلك ، إذا قمنا بتقييم النقاط التي لا نثق فيها إلا بالقيمة المتوقعة ، على الرغم من أننا تعلمنا المزيد عن المشهد ، فإن هذه التجارب باهظة الثمن في معظم الحالات تضيع على زيادة مستوى الثقة في المناطق منخفضة الأداء بدلاً من التركيز على إيجاد الحد الأقصى . وبالتالي ، إذا وجدنا نقطة جيدة ، فنحن نريد استغلال هذه النتيجة للبحث في مكان قريب عن حل أفضل (مع توقع أكبر).
يتم اقتراح عدة خوارزميات لموازنة المفاضلة بين الاستكشاف والاستغلال وتمثل أقصى وظيفة الاستحواذ مفاضلة تلقائية بين هذين العاملين. واحدة من وظائف الاستحواذ الشائعة الاستخدام هي “التحسين المتوقع” (EI) حيث تقوم الخوارزمية بتقدير مقدار التحسن المتوقع على أفضل ما هو متوقع من كل نقطة من النقاط ، وعينات النقطة بأعلى تحسن متوقع. تعمل هذه الوظيفة على إيجاد التوازن في الاستكشاف والاستغلال بمفردها باستخدام GP المدربة بالفعل وتجد النقطة التي توفر أعلى مستوى من التحسن المتوقع وتم اختيارها كدالة اقتناء في هذا العمل.
كما هو موضح سابقًا ، حسب التصميم ، يعتمد تحسين بايزي على تجارب متتابعة. في كل مرة يتم فيها تقييم نقطة واحدة ، يتم إعطاء النتيجة للخوارزمية لتحديث GP السابق والعثور على النقطة التالية التي سيتم تقييمها باستخدام وظيفة الاستحواذ. ومع ذلك ، يكون إجراء بعض التجارب أكثر فعالية بشكل متوازٍ وعلى دفعات متسلسلة ، وذلك لتقليل عدد جولات التجربة وبالتالي وقت المشروع بأكمله. لحسن الحظ ، تم تطوير مجموعة متنوعة من التحسينات البيزية مؤخرًا لتطبيقات المعالجة الموازية متعددة النواة. يمكن لهذه الخوارزمية التعامل مع تقييمات متعددة معلقة ويمكن أن تحصل على نتيجة أي من التقييمات المعلقة في أي وقت معين وإرجاع النقطة التالية ليتم تقييمها. باختصار ، تأخذ الخوارزمية في الاعتبار النتائج المحتملة لكل نقطة من النقاط المعلقة وتحسب وظائف الاستحواذ بناءً على جميع النتائج الممكنة. تم استخدام هذه الطريقة لدفع اتجاه تجاربنا وتم اختيار وتقييم مجموعة واحدة من النقاط في كل جولة وتم إعطاء النتيجة للخوارزمية لإنشاء المجموعة التالية من النقاط التي سيتم تقييمها. تجدر الإشارة إلى أنه في الإعداد التجريبي وعندما يتم إجراء التقييمات باستخدام تجربة موازية ، يتم تحديث النقاط المعلقة في نفس الوقت على دفعات لاحقة وليس واحدة تلو الأخرى.
إذا لم يكن هناك خطأ في التجارب ، وهذا هو الحال بالنسبة لتقييم الوظائف الرياضية ، فسيكون مستوى الثقة حول النقاط التي تم تقييمها بالفعل مرتفعًا للغاية. ومع ذلك ، نظرًا لأن نتيجة جميع التجارب تحتوي على بعض الأخطاء وهي أبعد ما تكون عن الحسابات الرياضية المثالية ، فقد تم تعديل الثقة في النتائج وبالتالي يتوقع البرنامج حدوث خطأ في التقييمات ويعدل المتوسط والاختلاف لجميع النقاط وفقًا لذلك. يتم شرح الجوانب الأخرى لخوارزمية التحسين هذه ، بما في ذلك وظائف التغاير ومعلمات فرط GP في التفاصيل بواسطة Sneok وزملاء العمل .
تقييم خوارزمية التحسين بايزي
لتوضيح التحسين بايزي مع GP ، قمنا بتعريف دالة واحدة متغيرة وحاولنا العثور على الحد الأقصى للقيمة عن طريق أخذ عينات متتابعة (الشكل ). تم اختيار الوظيفة عن عمد لتكون لها قمم متعددة و optima محلي (منحنى متقطع في الشكل) لاختبار ما إذا كانت خوارزمية التحسين يمكنها بالفعل العثور على الحد الأقصى العام. تمكنت الخوارزمية من العثور على الحد الأقصى ويتم توضيح مفاضلة الاستكشاف والاستغلال بواسطة ترتيب أخذ العينات المبين في الشكل. كلما زاد عدد النقاط التي تم تقييمها بواسطة الخوارزمية ، أصبحت الخوارزمية أقرب إلى الحد الأقصى كما هو موضح في الشكل .
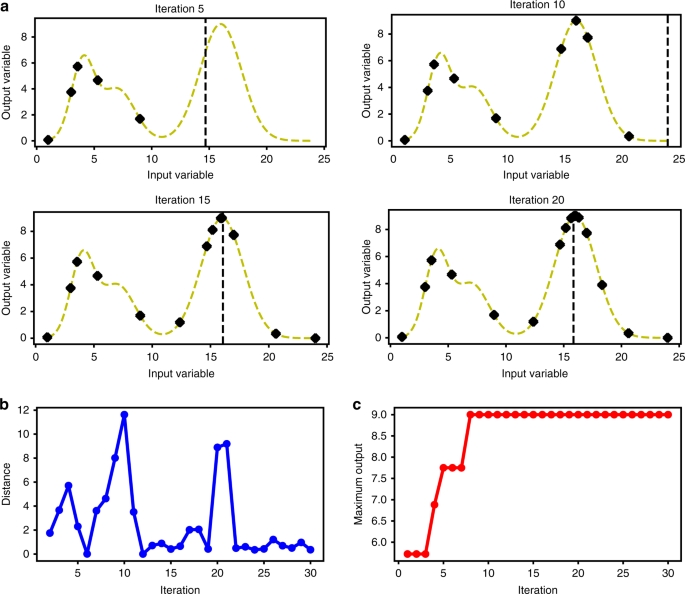
اختبار تحسين Bayesian من خلال إيجاد الحد الأقصى لوظيفة ثنائية الأبعاد. تقرر وظيفة الاستحواذ المدخلات التالية للاختبار ويتم استخدام الإخراج لتحسين النموذج التنبئي. يتم عرض التكرار 5 و 10 و 15 و 20 من هذه العملية. (ب) مع زيادة جولات التكرار ، يزداد النموذج التنبؤي ثقةً في موقع الحد الأقصى العالمي وتقل المسافة بين المدخلات المختبرة مع كل تكرار. ج تقييم الخوارزمية 9 نقاط قبل العثور على موقع الحد الأقصى. التكرارات اللاحقة ضبطت هذا التقريب نحو الأمثل الحقيقي. خوارزمية تقييم 12 نقطة قبل العثور على الحد الأقصى. يظهر الترتيب الذي يتم به تقييم كل نقطة على الرسم البياني
سعينا بعد ذلك إلى توضيح طريقة التحسين من خلال وظيفة مشابهة ذات 3 متغيرات مع ثلاثة مدخلات ومخرجات واحدة لمحاكاة مشكلة تحسين متعددة الأبعاد مماثلة. من الجدير بالذكر أنه تم استخدام التحسين البايزي في العديد من التطبيقات والغرض من هذه المحاكاة هو اختبار الخوارزمية على إعداد بسيط ولكن مماثل. تم ضبط محيط البحث على 1-24 لكل من المدخلات وتم تعيين الحد الأقصى للوظيفة على 9 ( y = f ( x 1 ، x 2 ، x 3 ) | x i ɛ {1، 2، … ، 24} ، f max = 9). تمكنت خوارزمية تحسين بايزي من العثور على الحد الأقصى لقيمة هذه الوظيفة من خلال تقييم 12 نقطة فقط من إجمالي 24 3 = 13،824 نقطة. كانت هذه التقييمات الـ 12 نتيجة لتكرار 12 اختبارًا واختبارًا ، مع اتباع كل تقييم خطوة تعلم أنتجت النقطة التالية للتقييم. سعينا بعد ذلك إلى مقارنة استراتيجية التحسين هذه بنهج أساسي حيث يتم تقييم النقاط التي تم أخذ عينات منها عشوائيًا ويتم استخدام كل هذه النقاط لتدريب نموذج الانحدار الذي يستند إلى تقدير المشتق الخارجي (EDE) الموضح في المنشورات السابقة. لقد وجدنا أنه على الرغم من أن أسلوب EDE يظهر قدرة تنبؤية مثيرة للإعجاب ، خاصة بالنظر إلى أن جميع البيانات قد تم الحصول عليها دفعة واحدة وليس من خلال أخذ العينات التكرارية ، حتى بعد أخذ 192 نقطة عشوائية ، لم يمكن العثور على الحد الأقصى (الجدول التكميلي).
ثم قمنا باختبار طريقة التحسين Bayesian عن طريق تشغيل عمليات محاكاة متعددة مع ظروف مختلفة.أولاً ، لمعرفة ما إذا كان بإمكان الخوارزمية العثور على الحد الأقصى من الوظائف الأخرى غير تلك التي تم اختبارها في القسم السابق ، أنشأنا 100 نموذج خليط غاوسي عشوائيًا ووجدنا الحد الأقصى لكل هذه الخوارزميات باستخدام هذه الخوارزمية. في المتوسط ، استغرق تقييم الخوارزمية 9.82 و 7.93 للعثور على الحد الأقصى و 95٪ من الحد الأقصى ، على التوالي. لاختبار تأثير الخطأ على الخوارزمية ، اخترنا بشكل عشوائي أحد نماذج الخليط الجاوسية 100 هذه وحاولنا إيجاد الحد الأقصى مع إضافة معدل الخطأ 0٪ و 10٪ و 20٪ ، الحد الأعلى لمعظم الطرق التحليلية ، إلى المخرجات قيمة تقييم الوظيفة لمحاكاة الإعداد التجريبي الحقيقي بشكل أفضل. لاحظنا أن الخوارزمية لا تزال قادرة على العثور على الحد الأقصى للوظيفة في معظم عمليات التشغيل ، ولكن زاد عدد التقييمات في كل عملية تشغيل بشكل كبير وفي حالة حدوث خطأ بنسبة 20٪ ، لم تتمكن من العثور على الحد الأقصى لـ 18٪ من الحالات حتى بعد 400 التقييمات. ومع ذلك ، يمكن أن تجد الخوارزمية 95 ٪ من الحد الأقصى في جميع الحالات (الجدول). هذا يدل على أن الخطأ ، كما هو متوقع ، يجعل التحسين أكثر صعوبة ، لكن خوارزمية تحسين Bayesian يمكنها ضبطه وتجد الحد الأقصى لمعظم الحالات. تجدر الإشارة إلى أن العثور على الحد الأقصى يزداد صعوبة مع ارتفاع معدل الخطأ. بخلاف حقيقة أن البيانات منخفضة الجودة ، كما هو متوقع ، تقلل من القدرة التنبؤية للنموذج ، مع ارتفاع معدل الخطأ ، يصبح الفرق بين النقاط الأقرب إلى الحد الأقصى غير قابل للتمييز.
أخيرًا ، تم تعييننا لتحسين عدد النقاط التي تم تقييمها في كل جولة ، مع المفاضلة بين التكلفة التجريبية والوقت: مع زيادة حجم كل دفعة ، تزداد تكلفة التجربة أيضًا ، ومع ذلك ، يزيد عدد الجولات الإجمالية من التجربة ، وبالتالي يقلل من الوقت الذي يقضيه في المشروع بأكمله. يتم تقييد أحجام الدُفعات أيضًا بالظروف التجريبية خاصةً بالنظر إلى التنسيق القياسي 96-جيد للتجارب البيولوجية عالية الإنتاجية. تم محاكاة عدد قليل من أحجام الدُفعات في نموذج الاختبار الموضح أعلاه مع تضمين نسبة الخطأ 10٪. لقد وجد أن أحجام الدُفعات التي يزيد حجمها عن 46 لم تقلل بشكل كبير عدد الدورات في مخطط التحسين ثلاثي الأبعاد (جدول تكميلي 2 ) وتم اختيار 46 كحجم الدُفعة لتجارب تحسين المسار في هذا العمل.
التحسين الآلي للمسار الحيوي اللايكوبين
بعد وضع اللمسات الأخيرة على نموذج التنبؤية وسياسة الاستحواذ ، اخترنا تحسين مسار الليكوبين الحيوي الاصطناعي كنظام نموذجي. أحد أسباب تدني الإنتاجية والإنتاجية في مسار التخليق الحيوي هو عدم توازن التدفق حيث تؤدي معدلات التفاعلات دون المستوى الأمثل إلى تراكم أو نضوب الجزيئات الوسيطة في التفاعل. هذا مهم بشكل خاص في المسارات ذات التفاعلات المتعددة حيث يصعب العثور على التوازن المعقد لكل خطوة من المسار. لقد ثبت أن ضبط تدفق كل خطوة في المسار وتحسينه يمثل استراتيجية فعالة للغاية لزيادة التدفق الكلي في مجموعة متنوعة من الحالات المختلفة. يمكن تمثيل التجريد من هذه المشكلة بمشهد إنتاج-التعبير حيث يتحقق أقصى تدفق من خلال مستوى تعبير معين لكل من الجينات في المسار. يجب علينا بعد ذلك تصميم إعداد تجريبي حيث يمكننا ضبط تعبير الجينات في المسار (مدخلات الوظيفة) وتحديد المخرجات التي نريد تعظيمها. يجب علينا بعد ذلك تجربة مستويات تعبير مختلفة كمدخلات والحصول على إنتاج اللايكوبين كإخراج والعثور على المدخلات التي تتوافق مع أعلى ناتج.
لأداء توليف التعبير لتحسين المسار وتوليد المدخلات ، يجب تطوير مجموعة من العناصر التنظيمية للتحكم في مستوى تعبير الإنزيمات في مسار الاهتمام. بالاعتماد على الأعمال المنشورة سابقًا ، قمنا بتحويل منطقة في مُروج T7 والتي تُعزى إلى قوتها لبناء 12 مُروجًا بمستويات تعبير مميزة. ثم صممنا واختبرنا موقعين لربط الريبوسوم (RBS) باستخدام حاسبة مكتبة RBS مع نقاط قوة مختلفة إلى حد كبير.أسفر مزيج T7p-RBS الناتج عن 24 مستوى تعبير متميز (الشكل التكميلي) مع مدى ديناميكي يبلغ 1000 ضعف. لاستكشاف ما إذا كان اتجاه مستوى التعبير الذي تم قياسه باستخدام eGFP يترجم إلى الاتجاه مع جينات crtE و crtB و crtI المتلقية للمعلومات من RBS ، تم دمج هذه الجينات مع eGFP ولكل من الجينات ، أربع مجموعات مروج / RBS من أربع مجموعات مميزة من المروج ضعيفة / قوية / RBS ، وتمت مقارنة كل اختار عشوائيا من قوة واحدة أو مستوى التعبير التعبير ، ولوحظ نفس الاتجاه التعبير العام (الشكل التكميلي 3 ).
تم تنفيذ سير عمل تحسين المسار باستخدام iBioFAB والذي تم استخدامه لتوليف TALEN عالي الإنتاجية وهندسة جينوم الخميرة الآلية . من خلال تسخير قوة iBioFAB وكذلك تحسين Bayesian، تم أتمتة جميع جوانب دورة DBTL. في كل جولة ، اختارت خوارزمية تحسين Bayesian 46 نقطة ليتم تقييمها (الرقم الذي تم اختياره بناءً على الاختبارات المذكورة أعلاه) ومنحهم برنامج جدولة iBioFAB. كعنصر تحكم ومحاسبة عن أي اختلافات بين دفعات مختلفة ، كانت واحدة من النقاط المختارة دائما النقطة الوسطى (12 ، 12 ، 12). قام البرنامج بعد ذلك بتمديد الأجزاء الصحيحة ليتم تجميعها من مكتبة الأجزاء وتجميع البلازميدات باستخدام مجموعة Golden Gate. تم بعد ذلك قياس إنتاج اللايكوبين للنقاط بأربع مكررات بيولوجية وتم إعادة القيم المتوسطة للنتائج إلى الخوارزمية لحساب النقاط التالية التي سيتم تقييمها. تبدأ خوارزمية التحسين النظرية الافتراضية باستكشاف المناظر الطبيعية بالكامل (الشكل) وتصبح أقل اتساقًا في الجولات اللاحقة (الشكل) حيث تتوفر المزيد من المعلومات حول المناظر الطبيعية ، مما يؤدي إلى استكشاف مناطق معينة. في الجولة 2 ، لا يزال هناك بعض الاستكشاف بينما تقارب النقاط في الجولة الثالثة تقريبًا إلى منطقة واحدة محددة يُعتقد أنها تحقق أعلى إنتاج لليكوبين.
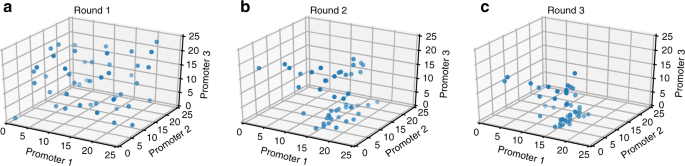
التغير في سلوك أخذ العينات من Baysesian الأمثل لمسار إنتاج اللايكوبين. في الجولة الأولى ( أ ) ، تم اختيار جميع النقاط لاستكشاف المناظر الطبيعية بشكل موحد لأنها غير مستكشفة وغير معروفة تمامًا ( ن = 46). في الجولة الثانية ( ب ) ، يتم الحصول على بعض المعلومات وتكون النقاط المنتقاة بواسطة الخوارزمية مائلة بشكل واضح من التوزيع الموحد ( ن = 45). ومع ذلك ، نظرًا لوجود بعض عدم اليقين ، فإنه لا يزال استكشاف المشهد.أخيرًا ، في الجولة الثالثة ( ج ) ، لوحظ وجود نمط واضح حيث حددت الخوارزمية أن النقاط في منطقة معينة تكون أكثر احتمالًا لتكون أقرب إلى أوبتيما العالمية وتستكشف هذه المنطقة بنشاط ولكن لا تزال تقوم ببعض التنقيب عن الحد الأدنى ( ن = 45). يتم توفير بيانات المصدر كملف بيانات المصدر
تتم مقارنة توزيعات إنتاج اللايكوبين بين النقاط التي تم تقييمها في كل جولة مع بعضها البعض في الشكل 4، ويلاحظ أن الجولات الأخيرة من تحسين المسار لها إنتاج أعلى متوسط للليكوبين وإنتاج أقصى أعلى مما يدل على فعالية خوارزمية تحسين بايزي في العثور على نقاط أفضل (أي المسوخ) في كل جولة لاحقة.لمقارنة خوارزمية تحسين بايزي بشكل أفضل مع عرض المكتبة العشوائية التقليدية ، تم إنشاء مكتبة عشوائية و 46 (نفس عدد نقاط كل جولة) و 136 (نفس عدد عدد النقاط في الجولات الثلاثة) بشكل عشوائي تم قياس إنتاج اللايكوبين الذي تم انتقاؤه ، كما يظهر الشكل 4 توزيعات إنتاج هاتين المجموعتين. يبلغ متوسط وأقصى عيار اللايكوبين الذي يتم العثور عليه عن طريق الفحص العشوائي 1.43 و 1.93 مرة أقل من تلك الموجودة في الجولة الثالثة من تحسين المسار. حتى من خلال تقييم 136 نقطة عشوائية ، كان الحد الأقصى لمقياس اللايكوبين الموجود أقل 1.77 مرة من الحد الأقصى من الجولة الثالثة.
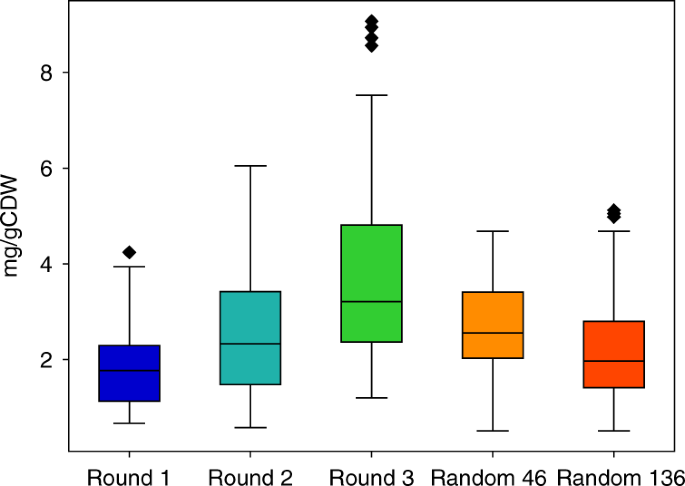
إنتاج الليكوبين في جولات مختلفة من تحسين المسار والفحص العشوائي. زادت النقاط المتوسطة والحد الأقصى بعد كل جولة من تحسين المسار. علاوة على ذلك ، على الرغم من أن متوسط وأقصى تقييم لـ 46 و 136 نقطة عشوائية يزيدان قليلاً عن التوزيع الموحد في الجولة 1 ، إلا أنهما أقل بكثير من النقاط التي اختارتها الخوارزمية في الجولات اللاحقة. تحتوي مربعات المخططات على بيانات داخل النطاق الرباعي (IQR) ، بينما تنتشر الشعيرات من الصناديق إلى 1.5 ضعف معدل الذكاء. خط الوسط في المربعات هو وسيط البيانات ونقاط أعلى الشعيرات هي قيم أعلى من معدل ضرب معدل الذكاء بمقدار 1.5 مرة أعلى من الربع الثالث. ن = 46 ، 45 ، 45 ، 46 و 136 لكل قطعة ، على التوالي. يتم توفير بيانات المصدر كملف بيانات المصدر
لمقارنة أفضل لأخذ العينات العشوائية مع تحسين بايزي ولتمثيل أكثر موثوقية الحد الأقصى الذي تم العثور عليه بواسطة أخذ العينات العشوائية ، تم تصميم نموذج توزيع على أساس 136 نقطة تم اختبارها بشكل عشوائي. أولاً ، تم حساب المتوسط والانحراف المعياري للبيانات التجريبية واستخدامها لإنشاء توزيع طبيعي. تم اختيار 136 نقطة بشكل عشوائي من هذا التوزيع وتم تسجيل الحد الأقصى. تكرر هذا 1000 مرة ويظهر التوزيع بين الحد الأقصى في الشكل التكميلي. تم العثور على المتوسط والانحراف المعياري لمجموعة بيانات 1000 maxima لتكون 4.81 و 0.43 ، على التوالي ، وبالتالي فإن النتيجة المتوقعة لأفضل متحولة من أخذ العينات العشوائية 136 نقطة هي 4.81 ± 0.43. هذه المحاكاة بعيدة عن الكمال لأن التوزيع الطبيعي ليس بالضرورة أفضل تمثيل للمناظر الطبيعية ، ولأن العبء الاستقلابي ربما يكون قد قلل من متوسط كمية الإنتاج من المسوخات التي تم اختيارها عشوائيًا. ومع ذلك ، فإنه يوفر خط أساس مفيد للمقارنة. كان أقصى 136 نقطة تم اختبارها في تجربتنا 5.12 ، ضمن النتيجة المتوقعة المحسوبة. هذا النطاق أقل بكثير من 9.07 ، أفضل متحولة تم العثور عليها باستخدام طريقة بايز الأمثل.
تم عزل أفضل منتجي اللايكوبين في كل جولة وميزوا في أنابيب الاختبار ، وتم تقدير كمية اللايكوبين باستخدام طريقة استخراج الأسيتون التقليدية. تم اختيار المسار ذي التعبير المتوسط المستوى لجميع الجينات كعنصر تحكم ، وتم تحليل مستويات إنتاج اللايكوبين لجميع هذه العينات الأربع في دفعة واحدة مع أربع مكررات بيولوجية ومقارنتها مع التحكم (الشكل التكميلي). وقد لوحظ أن أفضل توليفة في كل جولة قد زادت بشكل ملحوظ وأن أفضل سلالة عامة لإنتاج الليكوبين أفضل بثمانية أضعاف من التحكم.
نقاش
في هذا العمل ، قدمنا منصة تحسين مؤتمتة بالكامل تعتمد على الخوارزمية لتصميم النظم الحيوية حيث يقوم الجهاز بتنفيذ جميع الخطوات في عملية التحسين. تم دمج iBioFAB مع خوارزمية التعلم الآلي ، حيث بعد التصميم والإعداد المبدئي ، تقرر الخوارزمية التجارب التي يجب إجراؤها، ويقوم الروبوت بإجراء التجارب ويعيد البيانات إلى الخوارزمية وسيحدد النقطة التالية التي سيتم تقييمها. يتيح التعلم الآلي استكشاف مشكلات التحسين الكبيرة الأبعاد في حين يقتصر الحدس لدينا على ثلاثة أبعاد. على وجه الخصوص ، يتيح التعلم الآلي تحسينًا أسرع وأكثر استهدافًا من خلال التركيز فقط على المجالات ذات الاهتمام الكبير وعدم اليقين ، ويتعامل مع البيانات التجريبية من خلال الحفاظ على عدم اليقين من التجارب في الاعتبار ، ويحاول بنشاط تقليل عدد التجارب والتكلفة. BioAutomata أقل تحيزًا ، ويمكنه معالجة البيانات عالية الأبعاد ، ويرتكب أخطاء أقل ويمكنه العثور على الأمثل مع عدد قليل جدًا من التقييمات.
لإظهار أحد تطبيقات BioAutomata وكدليل على المفهوم ، وضعنا لتحسين تدفق المسار الحيوي اللايكوبين. تمكنا من ضبط التعبير الجيني لمسار 3 الجينات هذا لإيجاد التعبير الأمثل لإنتاج معظم اللايكوبين من خلال تقييم <1 ٪ من جميع 13.824 الاحتمالات. لقد قارنا أيضًا مخطط التحسين هذا بنموذج انحدار آخر تم الإبلاغ عنه سابقًا وكذلك أخذ عينات عشوائية ووجدنا أنه أفضل من كلاهما في الأداء. أفضل متحولة تم العثور عليها باستخدام BioAutomata أنتجت مقياس لايكوبين أعلى بواقع 1.77 مرة من أفضل متحولة تم العثور عليها باستخدام أخذ العينات العشوائية والمحاكاة أظهرت أن عدد التقييمات كان أقل بثمان أضعاف من مخطط التحسين القائم على الانحدار. تركز التحسين الذي تم إجراؤه هنا على المعلمات الجوهرية للمسار. من خلال تحسين المعلمات الخارجية ، مثل التحكم في التدفق عن طريق حذف الجينات التي تعتمد على المسار أو الإفراط في التعبير عن الجينات التي تتغذى على المسار ، أو هندسة الأيض المركزي ، أو تحسين الإجهاد أو تحسين التخمير ، تم الإبلاغ عن التتر العالي من تعبير اللايكوبين في الأدبيات.
تم اختيار المسار الحيوي لليكوبين في هذه التجربة على وجه التحديد بسبب طرقه الواضحة في الاستخراج والتقدير التي سهلت تنفيذ إنتاجية عالية باستخدام المعالجة الحيوية الآلية في ذلك الوقت. تشمل التحديات المحتملة للتطبيق العالمي لـ BioAutomata لتحسين المسار طرق استخراج يصعب تنفيذها على منصة آلية ، أو طرق تحليلية / كميّة تتطلب معدات أكثر تعقيدًا من قارئ اللوحة ، مثل مطياف الكتلة اللوني للغاز (GC-MS ) أو أجهزة القياس الكروماتوجرافي السائلة (LC-MS) السائلة. يمكن التغلب على هذه التحديات ، ولكن يجب بناء قاعدة حيوية واسعة النطاق ومتطورة لدمج هذه الأدوات. تجدر الإشارة أيضًا إلى أن توصيف المُروج في هذا العمل تم إجراؤه بواسطة جين البروتين الفلوري الأخضر (GFP) وليس الجينات الحيوية اللايكوبين ، وعلى الرغم من أن هذا الاختبار هو وسيلة تستخدم على نطاق واسع لتوصيف المروج / RBS ، ، لم نقم بقياس مستوى تعبير البروتين لجينات crtE و crtB و crtI في المسار الحيوي لليكوبين والذي قد يؤدي إلى رسم خرائط أكثر دقة لمستوى تعبير المروج / RBS ومستوى التعبير.
على الرغم من أن الخوارزمية قوية بشكل خاص عند استخدامها مع نظام مؤتمت بالكامل مثل iBioFAB ، يمكن تبنيها بسهولة للاستخدام في الإعدادات شبه الآلية أو اليدوية حيث يكون تقليل عدد التقييمات أكثر أهمية بسبب التكلفة التجريبية الأعلى. علاوة على ذلك ، تتوفر نماذج أخرى وخوارزميات التحسين وتم اختيار GP بشكل رئيسي بسبب تنفيذه بنجاح في الأنظمة البيولوجية . مجال التحسين المحتمل هو التخمين الأولي للمشهد لتحسينه. هنا ، لم نقم بعمل أي افتراضات أولية حول المشهد ، ومع ذلك ، فقد تم استخدام النموذج المدرّب لنظام واحد كنقطة انطلاق لنظام مشابه ليكون وسيلة قوية ويمكن لهذا التخمين المتعلّم أن يؤدي إلى تقليل عدد التقييمات للعثور على الحد الأقصى . على سبيل المثال ، يمكن استخدام المؤخرة الخلفية المدربة على مسار مكون من 3 جينات كنقطة انطلاق لتحسين المسار نفسه بجينات إضافية لتحسينه.
يمكن استخدام هذا النهج و BioAutomata لمشاكل تحسين الصندوق الأسود الأخرى حيث تكون التقييمات صاخبة ومكلفة ولا تقتصر على تحسين المسار. أحد الأمثلة التي يمكن تصوره هو هندسة البروتين حيث يمكن إجراء تغييرات مختلفة على تسلسل البروتين باستخدام الأساليب التي وصفها روميرو وزملاء العمل أو باستخدام CRISPR المستندة إلى أدوات طفرة وتعديل نقطة فيفو ويوجد التغيير الأمثل باستخدام نهج مماثل. يمكن أيضًا استخدام سير عمل التحسين هذا في مناطق أخرى بدءًا من تحسين المخزن المؤقت والوسائط إلى هندسة الجينوم بحثًا عن أنماط ظاهرية مطلوبة. بالنظر إلى الطبيعة الفعالة للغاية لمجموعة Golden Gate المكونة من 4 قطع ، كان من المفترض أن جميع ردود الفعل قد نجحت ، والتي قد لا تكون افتراضًا صحيحًا في التجميعات أو أنظمة التحسين الأكثر تعقيدًا وخطوة مراقبة الجودة المضمنة وطريقة الكشف عن الخارج يجب أن تضاف لهذه النظم المعقدة والخطأ عرضة. افترضنا أيضًا نموذجًا موحدًا للضوضاء في منهج تحسين النظرية الافتراضية من أجل البساطة. على الرغم من أن نموذج الضوضاء هذا لا يتطابق مع النموذج المستخدم في تراجع GP النموذجي ، فقد أثبتنا أن طريقة التحسين Bayesian المستندة إلى GP هذه كانت قادرة على العمل بفعالية حتى مع عدم تطابق بعض النماذج المتواضعة. قد تجد التطبيقات المستقبلية التي يُتوقع فيها أن يكون تقلب ضوضاء القياس عبر التجارب مصدر قلق كبير ، من المفيد استخدام نماذج ضوضاء غير متجانسة .
إن احتمالية وجود أنظمة روبوتية قائمة على الخوارزمية في علم الأحياء الهندسي له العديد من الوعود والتحديات. من ناحية ، يعد الإشراف البشري أمرًا بالغ الأهمية للحفاظ على القضايا الأخلاقية المحيطة بالهندسة الذاتية للحياة ومراقبة مدى ما تقوم به الآلة وما تحققه. من ناحية أخرى ، يمكن لنظام روبوتي قائم على الخوارزمية مستقل ، مرتبط بشبكة المعرفة ، أن يتعلم من المعلومات المنشورة في الوقت الفعلي وينشر نتائج تجاربه في الوقت الفعلي أيضًا. بخلاف المزايا الواضحة المتمثلة في خفض التكلفة وزيادة دقة البحث ، يمكن لشبكة BioAutomata المتصلة تقليل الوقت بشكل كبير من إجراء التجارب إلى نشر البيانات واستخدامها من قبل الآخرين. ستستفيد BioAutomata استفادة كبيرة من توحيد البيانات والمعايير التالية التي وضعتها قواعد البيانات مثل قاعدة بيانات Braunschweig Enzyme (BRENDA) ، و موسوعة Kyoto of Genes and Genomes (KEGG) ، و Protein Data Bank (PDB) ، و Synthetic Biology Open Language (SBOL) .
طرق
سلالات زراعة
استخدمت DH5α و BL21 (DE3) Escherichia coli (نيو إنجلاند بيولاب ، إبسويتش ، MA) لصنع خلايا مؤهلة كيميائيا باستخدام مجموعة Mix & Go E. coli Transformation Kit (Zymo Research ، Irvine ، CA) لتضخيم البلازميد وإنتاج اللايكوبين ، على التوالي . نمت E. coli في وسط Luria Broth (LB) المتوسط (Fisher Scientific، Pittsburgh، PA) مستكمل بـ 50 ميكروغرام / مل من Spectinomycin (Spec) أو 25 ميكروغرام / مل كاناميسين (Kan) للحفاظ على البلازميد أو 0.5 مللي إيزوبروبيل – β – ث – ثيوكالاكتوزيد (IPTG) للتحريض حسب الاقتضاء. تم شراء المضادات الحيوية و IPTG من الذهب التكنولوجيا الحيوية (سانت لويس ، MO). نمت DH5α E. كولاي الخلايا و BL21 (DE3) الثقافات بداية الخلية في 37 درجة مئوية ، ولكن تزرع الثقافات خلية BL21 (DE3) لإنتاج الليكوبين في 28 درجة مئوية ، ودرجة حرارة النمو المثلى لإنتاج الليكوبين . تم حساب وزن الخلية الجافة (DCW) من OD 600 باستخدام dcw / OD من 0.36 كمعدل التحويل.
التلاعب الحمض النووي وبناء البلازميد
لتوليد متغيرات T7 (T7p) معزز بمستويات تعبير مختلفة ، تم تحوير المنطقة المنسوبة إلى قوتها 46باستخدام T7p-mut-3N و T7p-mut-6N ، عند تضخيم جين eGFP بأشكال T7t terminator (T7t). تم استنساخ جزء DNA T7p-mut-eGFP-T7t الناتج في العمود الفقري pET26 (ب) باستخدام ربط تقييد الهضم. ثم تم تحويل المكتبة الناتجة إلى خلايا المختصة BL21 (DE3) وتم انتقاء 192 مستعمرة بشكل عشوائي ونمت بين عشية وضحاها عند 37 درجة مئوية. في اليوم التالي ، تم تلقيح 900 ميكرولتر من LB + Kan مع 10 ميكرولتر من ثقافة البذور وحضنت عند 37 درجة مئوية و 250 دورة في الدقيقة. بعد 3 ساعات ، تمت إضافة 100 ميكرولتر من LB + Kan مع 5 مل IPTG إلى ثقافة الخلية وتم حضنها عند 28 درجة مئوية. بعد 4 ساعات ، تم قياس مضان eGFP (انبعاث 488 نانومتر / 509 نانومتر) ، وكذلك OD 600 ، وتم اختيار 24 مروجًا مختلفًا لمزيد من التوصيف. تم تصميم تسلسلين RBS باستخدام حاسبة مكتبة RBS لتنظيم الترجمة وتم دمجها مع المروجين الذين تم تحديدهم لتقييم قوتهم. ثم تم استخدام هؤلاء المروجين لاستنساخ كاسيت التعبير T7-mut-RBS-eGFP-T7t. تم اختيار اثني عشر من هؤلاء المروجين الذين أظهروا مجموعة واسعة من نقاط القوة كمكتبة مروج لتنظيم النسخ. لاختبار مستوى التعبير عن جينات اللايكوبين ، تم تضخيم جينات crtE و crtB و crtIPCR ودمجها مع جين eGFP ومجموعات مختلفة من المروج / RBS باستخدام تجميع Gibson. تم إجراء تعبير الانصهار لجينات crt و eGFP باستخدام رابط مرن (GGATCCGCTGGCTCCGCTGGGTTCTGGCGAATTC) الذي تم تحسينه لتعبير الانصهار GFP في E. coli . ثم تم التعبير عن T7_mut_RBS (ضعيفة / قوية) _Crt (E / B / I) _eGFP شرائط التعبير في أربع مكررات بيولوجية تتبع نفس البروتوكول على النحو الوارد أعلاه وتم قياس مضان.
تم استخدام QIAGEN Plasmid Mini Kit (QIAGEN ، فالنسيا ، كاليفورنيا) لعزل البلازميدات من خلايا E. coli و Zymoclean Gel DNA DNA Kit (Zymo Research، Irvine، CA) تم استخدامه لتنقية الهلام. تم شراء جميع إنزيمات التقييد ، بوليميريز Q5 ، مكونات المزيج الرئيسي من Gibson Assembly وناقلات المكوك E. coli من New England Biolabs (Ipswich، MA) وتم شراء جميع المواد الكيميائية من Sigma-Aldrich (St. Louis، MO) ما لم يتم تحديد خلاف ذلك. يتم سرد جميع الاشعال والبلازميدات المستخدمة في هذه الدراسة في البيانات التكميلية، على التوالي. يتم تضمين ملفات GenBank مع خريطة مشروحة لأجزاء الحمض النووي وكذلك البنى النهائية في المعلومات التكميلية. تتوفر السلالات والبلازميدات من خلال اتفاقية نقل المواد القياسية من جامعة إلينوي.
جولدن جيت التجمع
تم استخدام طريقة تجميع البوابة الذهبية لتجميع مسار اللايكوبين بمستوى تعبير مختلف لكل جين. أولاً ، تم هضم بلازميد pSPE باستخدام إنزيمات تقييد AFI II و Xba I. بعد الهضم ، تم تلطيخ قليتين مكملتين تحتويان على جولدن جيت المحسنة، بالإضافة إلى مروج T7 ، فاصل وموقع التعرف على EcoR V ، تم تفسيرهما واستنساخهما بين مواقع القطع. تم إجراء الفسفرة باستخدام T4 Polynucleotide Kinase (New England Biolabs، Ipswich، MA) ، باتباع إرشادات الشركة المصنعة. تم تضخيم البلازميد بعد ذلك ، وتم هضمه باستخدام EcoR V ، وتم استنساخ كل من جينات crtE و crtB و crtI بنقاط قوة معزز مختلفة لـ RBS / T7 باستخدام طريقة التجميع Gibson مع مروج T7 والمنفذ كأذرع متجانسة بواسطة NEBuilder HiFi DNA Assembly Cloning Kit (New England BioLabs، Ipswich، MA) كما هو موضح في البيانات التكميلية . لإنشاء إدراج للاستنساخ في البلازميدات المساعدة ، تمت إضافة RBS إلى كل من جينات crtE و crtB و crtI باستخدام تضخيم PCR وأضيف مروج T7 في خطوة أخرى من تفاعل PCR. ثم تم تضخيم 72 البلازميدات المجمعة في E. coli وتم تأكيد الإضافات بواسطة تضخيم PCR. تم الحصول على البلازميد pET26b من EMD Millipore (Billerica، MA) واستخدم كمستقبل لمسار الليكوبين الحيوي. تم وضع روابط Golden Gate ومواقع Bsa I على اثنين من ألكليوكليوتيدات تكميلية مما أدى إلى شظية قصيرة من الحمض النووي مع نهاية لزجة بعد الصلب والتفسخ. تم هضم بلازميد pET26b باستخدام إنزيمات Xho I و Sph I وربطها بجزء الحمض النووي الذي يحتوي على أجزاء متراكبة لبناء العمود الفقري لمسار إنتاج اللايكوبين.
تم تضخيم الأجزاء 73 التي تم تجميعها (72 إدراج و 1 عمود فقري) في E. coli وتنقيتها. تم ضبط تركيز العمود الفقري على 30 نانوغرام / ميكرولتر وتم ضبط تركيز بقية الأجزاء على نفس التركيز المولي.يتكون كل رد فعل من البوابة الذهبية 20 ميكرولتر من 100 نانوغرام من العمود الفقري وكميات متساوية الأقطاب من crtE و crtB و crtI ، و 10 وحدات من إنزيم تقييد Bsa I ، و 100 وحدة من Ligase DNA T4 ، و 2 ميكروليتر من عازلة CutSmart ، و 0.75 ميكروليتر من أدينوساين ثلاثي الفوسفات ( ATP) (25 مم). بعد رد فعل البوابة الذهبية ، 5 ميكرولتر من مزيج سيد نوكلياز يتكون من 2.5 وحدة من BSA I ، 2.5 وحدة من نوكلياز آمن البلازميد (Illumina ، سان دييغو ، كاليفورنيا) ، 0.5 ميكرولتر من CutSmart العازلة و 1 ميكرولتر من ATP (25 ملي مولتر) تمت إضافة إلى رد الفعل على خطي أي العمود الفقري عسر الهضم وهضم جميع الأجزاء الخطية من الخليط. لقد تم اعتماد البوابة الذهبية المذكورة أعلاه وبروتوكول المزيج الرئيسي الآمن للبلازميد من عملنا السابق مع بعض التعديلات ولكن بروتوكول التدوير الحراري لم يتغير. لضمان التجميعات عالية الكفاءة ، تم اختيار روابط Golden Gate المحسنة لهذه التجربة من مجموعة عالية الكفاءة من الروابط . لاختبار كفاءة وإخلاص مجموعة Golden Gate ، تم إجراء 24 تفاعلًا باستخدام كلٍ من الأجزاء الـ 72 مرة واحدة على الأقل وتم اختيار أربعة مستعمرات من محولات كل رد فعل وتم تأكيد جميع التجميعات على أنها صحيحة. يتم عرض عينة من منتجات التجميع هذه في البيانات التكميلية .
استخراج الليكوبين والكمي
يمكن استخراج اللايكوبين بواسطة المذيبات العضوية والكمية المسعرية عن طريق قياس الامتصاصية في حوالي 470 نانومتر. هذا الاختبار حساس للغاية وتم الإبلاغ عن تحديد كمية اللايكوبين بدقة المليغرام الفرعية. الطريقة الأكثر شيوعًا لاستخراج الليكوبين وتقديره تنطوي على إعادة تعليق الخلايا في الأسيتون تليها الحضانة في الأسيتون . نظرًا لأن الأسيتون شديد التقلب ويذيب أختام الغراء وبعض المواد الاستهلاكية الأخرى ، فهو ليس مثاليًا للاستخدام في نظام التشغيل الآلي. لذلك ، تم اختبار أربعة مذيبات عضوية أخرى، بعضها في المنشورات السابقة، للتأكد من فعاليتها في استخراج اللايكوبين. تم العثور على أكثر مذيبات الاستخراج فعالية والذي يتوافق مع أنظمة الإنتاجية العالية ليكون ثنائي ميثيل سلفوكسيد (DMSO). تم نسج خلايا E. coli وتم إزالة الطاف. ثم تم تعليق الخلايا في 300 ميكرولتر من DMSO وتم حضنها لمدة 30 دقيقة عند 37 درجة مئوية عند 250 دورة في الدقيقة. بعد الحضانة ، تم إزالة خليط الخلوي DMSO عند 3000 دورة في الدقيقة لمدة 10 دقائق وتم إزالة 200 ميكرولتر من طاف وتم قياس الامتصاصية عند 472 نانومتر وربطها بإنتاج اللايكوبين.
الأتمتة الكاملة لسير العمل
تم استخدام iBioFAB لأتمتة تجميع أجزاء الحمض النووي لمسار اللايكوبين ، والتحول ، وزراعة الخلايا واستخراج اللايكوبين. يوضح الشكل سير العمل الكلي للتجارب. أولاً ، يتم إنشاء الأجزاء التي سيتم تجميعها عن طريق خوارزمية التعلم الآلي ويتم إعطاؤها لمولد النص البرمجي الموصوف مسبقًا لتوليد مسارات pipetting لمعالج سائل Tecan. بعد ذلك تم لف ألواح خليط الحمض النووي ، واخلطها مع مزيج Golden Gate الرئيسي ونقلها إلى thermocycler لتفاعل Golden Gate. بعد 30 دورة من الهضم والربط في مجموعة Golden Gate ، تمت إضافة مزيج Master Plasmid Safe إلى المزيج تليها 30 دقيقة من الهضم باستخدام Bsa I و nuclease الآمن للبلازميد. بعد ذلك ، تم تحويل منتج تجميع Golden Gate الآمن المعالجة بالبلازميد في خلايا المختصة بالكولونية BL21 (DE3) E. و مطلي على ألواح أجار LB ونقله من على سطح السفينة للحضانة. تم تحضين الأطباق عند 37 درجة مئوية خلال الليل وتم اختيار أربع مستعمرات من كل لوحة باستخدام Pickolo colour-picker (SciRobotics، Israel) وتم تلقيحها في 1 مل من LB + Kan media. نمت ثقافة البذور بين عشية وضحاها وأضيفت 50 ميكرولتر من الثقافة إلى 800 ميكرولتر من وسائل الإعلام الطازجة LB + كان وحضنت في 37 درجة مئوية. بعد 2 ساعة ، تمت إضافة 200 ميكرولتر من LB + Kan + 2.5 مم IPTG إلى الثقافة وتم تحضين الخلايا المستحثة عند 28 درجة مئوية لمدة 24 ساعة لإنتاج أقصى .تم قياس OD 600 ثم تم تكوير الخلايا وإعادة تعليقها في DMSO وحضنت عند 37 درجة مئوية لمدة 30 دقيقة لتقدير الليكوبين. لتقليل الاختلافات المحتملة بين الجولة المختلفة من التحسين ، تم تكرار النقطة ذات مستوى التعبير المتوسط للجينات الثلاثة (12 ، 12 ، 12) في الجولتين الثانية والثالثة من التحسين. تم تضمين اثنين من الضوابط الأخرى OD (أي التلقيح والنمو) وإنتاج الليكوبين (البلازميد فارغة) في أربع مكررات في جميع الجولات الثلاث. لذلك ، كان العدد الإجمالي للنقاط الجديدة في الجولات الأولى والثانية والثالثة 46 و 45 و 45 ، على التوالي ، وكانت كل جولة تتألف من صفيحتين كاملتين تحتويان على 96 بئر. لاختبار كفاءة التجمع ، تم اختيار 24 من المجموعات المجمعة عشوائياً وتم التحقق منها من خلال هضم إنزيم مقيد وأثبتت جميعها 24 أنها صحيحة كما هو موضح في الشكل التكميلي. تم تسلسل ثلاثة من هذه البلازميدات الـ 24 أيضًا ، وكانت النتيجة مطابقة للتسلسل المتوقع (البيانات التكميلية ).
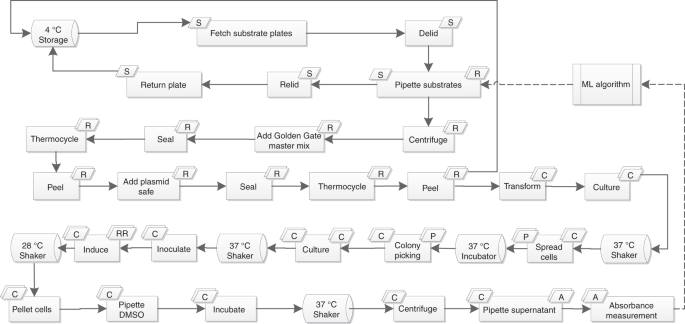
سير عمل تحسين المسار الكلي المؤتمت بالكامل. تقوم خوارزمية التعلم الآلي باختيار البلازميدات المراد تجميعها وإرجاع القائمة إلى iBioFAB لأداء التجميع. ثم يتم تحويل المنتجات المجمعة ، ويتم عزل أربعة مستعمرات مفردة لتقدير كمية اللايكوبين وقياس OD. ثم يتم إعطاء البيانات الناتجة إلى خوارزمية التعلم الآلي لاختيار المجموعة التالية من النقاط التي سيتم تقييمها.
 Loading...
Loading...